Overview
This is a four-part series covering gastrointestinal physiology.
- Part I – Overview of the gastrointestinal system & the oral cavity.
- Part II – Digestion: stomach & control of digestion.
- Part III – Secretion: pancreas & hepatobiliary system.
- Part IV – Absorption: small intestines.
In part III we look at the secretions of the hepatobiliary system and the exocrine pancreas. Their secretions, released into the small intestines are essential to the normal digestion and absorption of nutrients:
- Hepatobiliary system: this consists of the liver, gallbladder and bile ducts. Bile is secreted by hepatocytes into the biliary circulation. It is stored in the bile duct and released into the small intestines by the common bile duct which drains into the second part of the duodenum (via the ampulla of Vater).
- Exocrine pancreas: the exocrine pancreas produces amylase, lipase, proteolytic enzymes (e.g. trypsin) and an alkaline pancreatic juice which are released into the second part of the duodenum alongside the common bile duct via the pancreatic duct.
The pancreas
The pancreas is a retroperitoneal organ that has both exocrine and endocrine function.
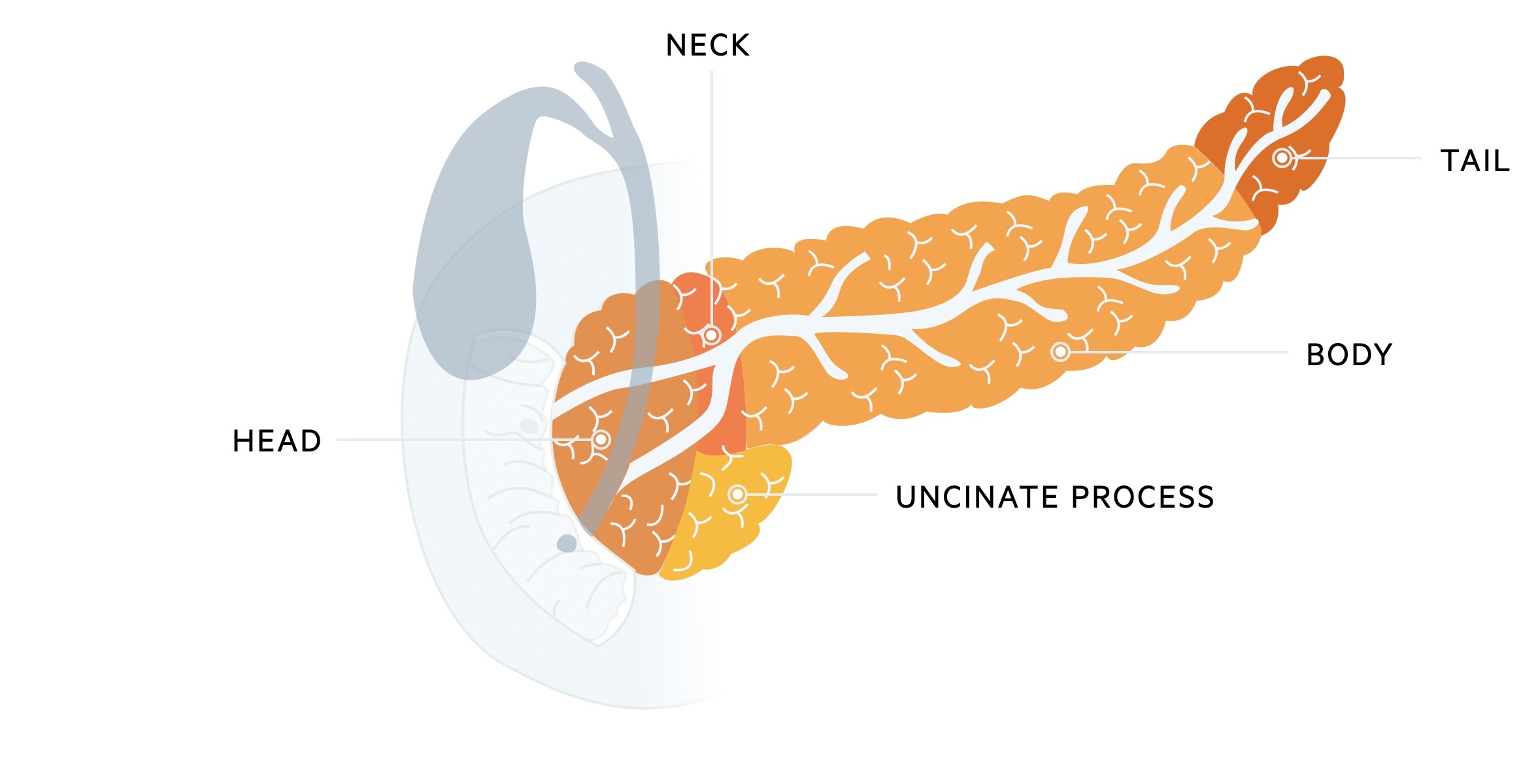
The pancreas is divided into five main regions; head, neck, uncinate process, body and tail. It first appears in the fifth week of gestation, developing from an outgrowth of the foregut. It develops as two separate buds (ventral and dorsal) which eventually fuse. The ventral bud forms the uncinate process and head of the pancreas whilst the dorsal bud forms the neck, body and tail of the pancreas. The duct associated with the ventral bud forms the main pancreatic duct (of Wirsung) whilst the duct associated with the dorsal bud forms the accessory duct (of Santorini).
Abnormal fusion between the ventral and dorsal buds lead to anatomical variations:
- Annular pancreas: incomplete rotation of the ventral bud leads to its surrounding of the 2nd part of the duodenum.
- Pancreas divisum: caused by failure of fusion of the ventral and dorsal buds. It is common, affecting around 4-15% of individuals
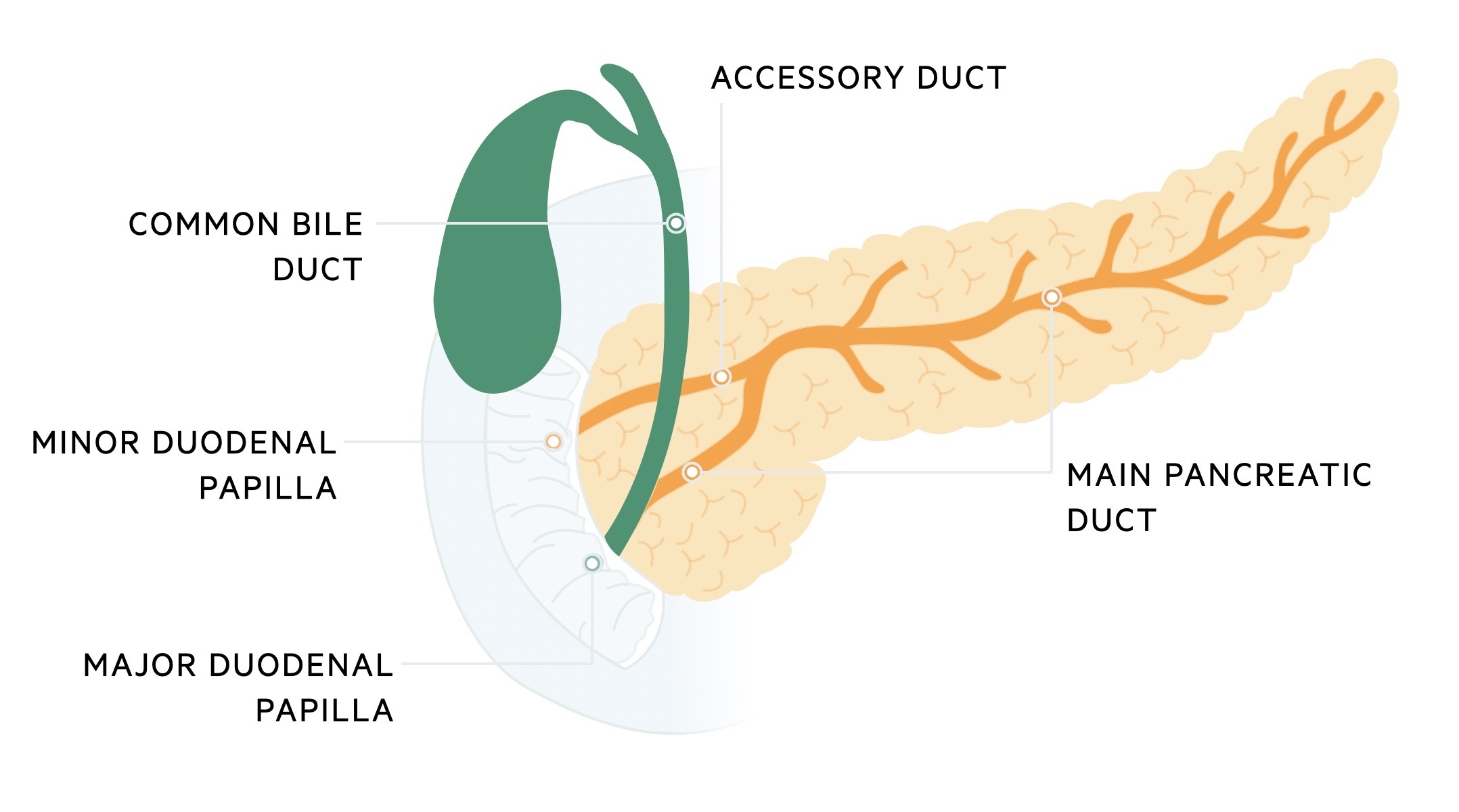
The secretory products of the exocrine pancreas are passed into the pancreatic ducts which drain into the duodenum. The pancreatic ducts exhibit a great deal of anatomical variability. The most common arrangement is a single main pancreatic duct and a single accessory duct which normally unite. The main pancreatic duct drains most of the pancreas, exiting into the major duodenal papilla in the second part of the duodenum via the ampulla of Vater The accessory duct is smaller and drains part of the pancreatic head, exiting via the minor duodenal papilla a few centimeters proximal to the major duodenal papilla.
Endocrine pancreas
Endocrine cells make up 1-2% of the pancreatic mass. These cells are organised into special structures called islets of Langerhans.
- Alpha cells: secrete glucagon, a hormone that opposes the actions of insulin.
- Beta cells: secrete insulin which stimulates cellular uptake of glucose and glycogen formation within the liver.
- Delta cells: secrete somatostatin an inhibitory hormone.
Exocrine pancreas
The exocrine pancreas accounts for approximately 98-99% of the pancreatic tissue. It is organised into secretory sacs termed acini. The ductal cells, which line the pancreatic ducts, are important in the secretion of a bicarbonate-rich alkaline solution. This helps to neutralise the acidic chyme as it arrives in the small intestinal. Approximately 1-2 litres of pancreatic secretions are produced each day.
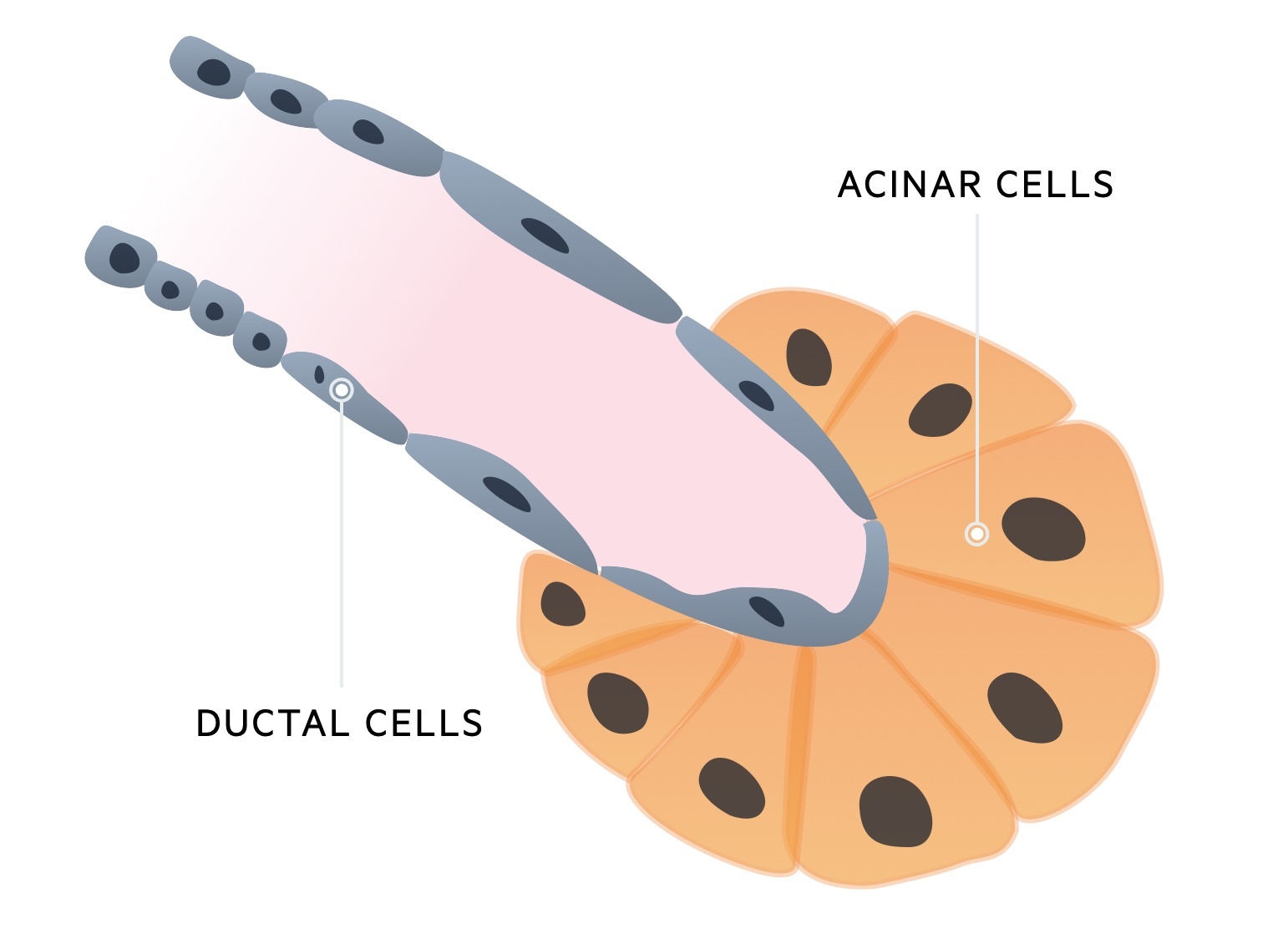
The acini drain to ducts which connect to form the main pancreatic duct that empties through the ampulla of Vater. These acinar cells are important into the secretion of pancreatic enzymes that help in the digestion of carbohydrates, fats and protein. Pancreatic enzymes work best in a neutral or slightly alkaline environment.
There are three important groups of enzymes that are secreted from the exocrine pancreas:
- Pancreatic amylase
- Pancreatic lipase
- Proteolytic enzymes
Pancreatic amylase is important for the digestion of carbohydrates. It is secreted in its active form and helps convert polysaccharides to the disaccharide maltose.
Pancreatic lipase is the only enzyme within the human body that is able to digest fats. It breaks down triglycerides into monoglycerides and free fatty acids. The action of pancreatic lipase is reliant on bile for the emulsification of fats. Like pancreatic amylase, it is secreted in its active form.
The pancreas secretes three main proteolytic enzymes; trypsinogen, chymotrypsinogen and procarboxypeptidase.
These enzymes are secreted in an inactive state to prevent proteolytic damage to the pancreas. The pancreas also secretes a trypsin inhibitor to further prevent proteolytic damage to its own cells.
Activation of these enzymes requires the enzyme enterokinase. Enterokinase (a luminal membrane enzyme) cleaves trypsinogen into trypsin rendering it active. Trypsin then cleaves chymotrypsinogen into chymotrypsin and procarboxypeptidase into carboxypeptidase.
Pancreatic insufficiency
The deficiency of pancreatic enzymes can lead to malabsorption.
There are many causes of malabsorption. The causes can be categorised based on the underlying pathological mechanism which include mucosal injury, pancreatic enzyme deficiency, bile deficiency, or lymphatic problems.
In the Western world, coeliac disease is one of the most common causes of malabsorption that results from dietary sensitivity to gluten. This leads to mucosal injury and, therefore, damage to the absorptive surface.
Some patients may develop pancreatic enzyme deficiency secondary to damage to the pancreas (e.g. chronic pancreatitis). These patients need pancreatic enzyme replacement to allow adequate digestion and absorption of nutrients.
Creon is a commonly used pancreatic supplement, which contains a mixture of digestive enzymes (e.g. lipases, proteases and amylases) from the pancreas of pigs.
The biliary system
The biliary system is composed of the gallbladder, the intrahepatic and extrahepatic ductal system.
The function of the biliary system is to transfer bile, produced by hepatocytes, into the duodenum to help with the emulsification of fats. Bile is continuously produced and secreted by hepatocytes into small channels called bile canaliculi. Bile is then transferred through the biliary system into the CBD and exits via the ampulla of Vater.
When bile is not required for digestion, it is stored within the gallbladder and can be excreted on the arrival of a fatty meal. Within the gallbladder, simple columnar epithelial cells are important in concentrating bile between meals. The gallbladder may hold up to 50mls of bile and concentrate it by 5-10 times.
The sphincter of Oddi guards the ampulla of Vater during times of fasting and prevents bile outflow into the duodenum. Cholangiocytes line the biliary ducts and they are important in the modification of bile as it is taken to the duodenum or stored in the gallbladder.
Bile
The primary function of bile is the emulsification of fats. It is also useful in the excretion of waste products.
Approximately 250-1000mls of bile is secreted each day. Bile has a number of important components including bile salts, bile pigments, lecithin (phospholipid), aqueous solution, cholesterol and inorganic salts.
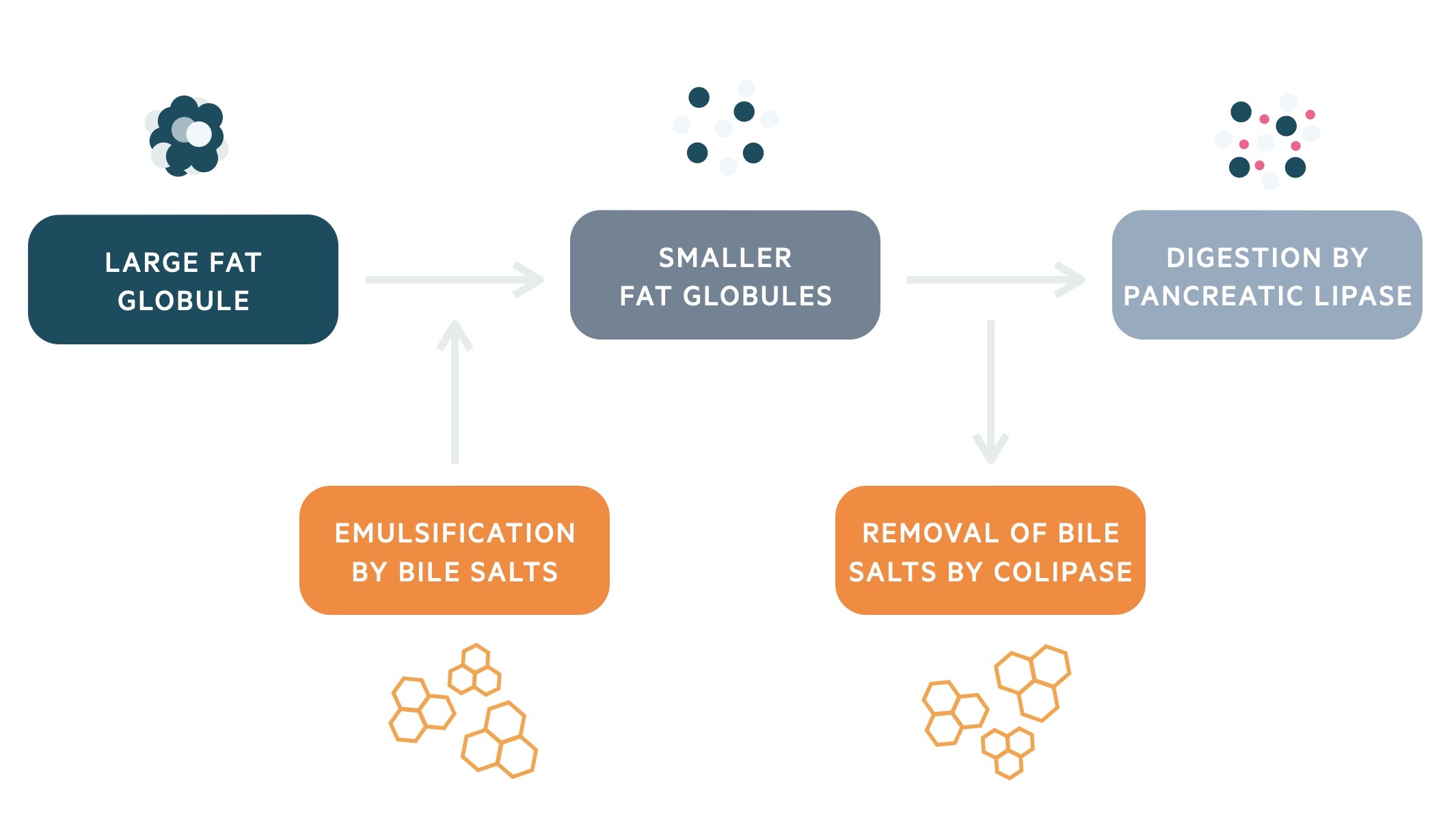
Dietary lipids are predominantly triglycerides. The hydrophobic nature of triglycerides means they clump together reducing the surface area. Bile salts help break up these large fat globules into smaller ones, thereby increasing the surface area through which pancreatic lipase may act.
Bile salts have a hydrophilic and hydrophobic component. As intestinal mixing breaks up the fat globules, the absorbed bile salts leave their hydrophilic portions on the outside. These hydrophilic portions repel one another (due to negative charge), which prevents the fat globule from reforming.
Finally, before pancreatic lipase can digest the fat globules its requires the action of an additional enzyme called colipase. Colipase is able to displace bile salts. It then binds to lipase and anchors it to the surface of the fat globule.
Digested lipids (including cholesterol and fat-soluble vitamins) are then aggregated into structures called micelles. Micelles have a lipid-rich core surrounded by a hydrophilic shell of bile salts and lecithin. Micelles, water-soluble structures, are taken to the luminal surface of enterocytes where the lipid content can be absorbed.
Bilirubin excretion
Bilirubin is the major excretory waste product of bile.
Bilirubin is a product of haemoglobin breakdown from within red blood cells (erythrocytes). The process of bilirubin excretion involves three key processes; release of bilirubin from haemoglobin, transfer to the liver for conjugation and excretion into bile.
Formation
Old erythrocytes can be broken down by macrophages predominantly located in the spleen. Other organs involved in this process include the liver and lymph nodes.
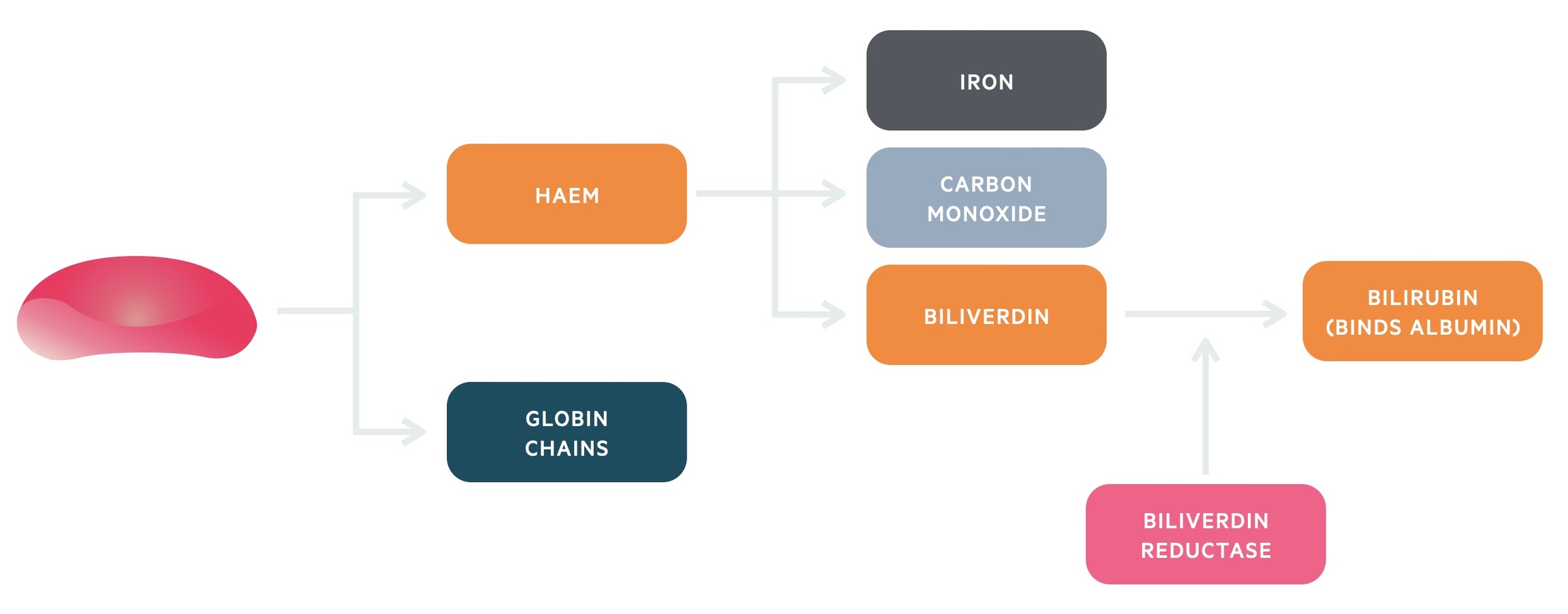
Initially, haemoglobin is broken down into haem and globin chains. The globin chains can be broken down into amino acids and recycled.
Haem is metabolised further by the enzyme haem oxygenase 1 (HO-1). This enzyme breaks haem into iron, carbon monoxide and biliverdin. Biliverdin is then reduced to bilirubin by the action of biliverdin reductase.
Transfer
Bilirubin is bound to albumin in the serum and carried to the liver. Within the liver bilirubin is conjugated by the enzyme glucuronyltransferase (full title: Uridine diphosphoglucuronate (UDP)-glucuronosyltransferases (UGTs)). This is a process that involves the addition of a glucuronic acid to bilirubin (glucuronidation). Reduced activity of this enzyme causes the condition Gilbert’s syndrome.
Gilbert’s syndrome, a common autosomal recessive condition, is caused by a mutation in the gene for UGT and impaired glucuronidation. It manifests as recurrent episodes of mild unconjugated hyperbilirubinaemia during times of intercurrent illness. During episodes, scleral jaundice can often be seen. Blood tests (except hyperbilirubinaemia) are usually normal and patients are asymptomatic with no treatment required.
Excretion
Once conjugated, bilirubin is excreted into bile and released into the duodenum. Bilirubin is converted into urobilinogen by bacteria. After conversion to urobilinogen, the excretion of bilirubin takes two different pathways: major and minor
The major pathway involves urobilinogen being converted by bacteria into stercobilin and urobilin. This remains within the intestines and is excreted within the faeces. These breakdown products give faeces its characteristic brown colour. The minor pathway involves some urobilinogen being reabsorbed into the circulation and then excreted by the kidneys. Excessive excretion of urobilinogen can lead to darkening of the urine.
In obstructive jaundice excretion of bilirubin via the major pathway is blocked. More bilirubin is excreted via the minor pathway through the kidneys. This leads to the characteristic clinical findings of pale stools and dark urine. Causes of obstructive jaundice may include carcinoma at the head of the pancreas or gallstone-related disease.
Control of hepatobiliary secretions
Control of hepatobiliary secretions occurs upon entry of acidic and fat-rich chyme into the duodenum.
The arrival of the acidic and fat-rich chyme leads to the release of two important enterogastrones: secretin and cholecystokinin (CCK). An enterogastrone is a hormone secreted by the upper intestinal mucosa that has an inhibitory action on gastric motility and secretion.
Secretin is released in response to acid in the duodenal lumen, where CCK is released in response to fat and protein. Both of these hormones have trophic effects (maintains the integrity of tissue) on their target organs (e.g. exocrine pancreas, gallbladder).
Role of secretin
Entry of acidic chyme stimulates S cells within the duodenal mucosa to secrete secretin. Secretin stimulates pancreatic ductal cells to secrete the an alkaline fluid, which is rich in bicarbonate ions. It also stimulates the bile duct epithelium to secrete an alkaline rich fluid. Collectively, this helps to neutralise the acidic chyme that enters the duodenum.
The amount of bicarbonate that is secreted is proportional to the amount of acid that enters the duodenum.
Role of CCK
Entry of fat and protein products of digestion stimulates I cells within the duodenal mucosa to secrete CCK. CCK stimulates the pancreatic acinar cells to secrete digestive enzymes into the pancreatic duct.
All three major pancreatic enzymes are packaged into zymogen granules and released together by exocytosis on stimulation. The total amount of enzymes released varies depending on the type of meal (fatty meals having the greatest effect), but the proportion of individual enzymes does not alter with meals. The content of the zymogen granules (e.g. proportion of different enzymes) can be altered long-term in accordance to changes in diet.
CCK also has an effect on the gallbladder. It initiates contraction of the gallbladder leading to the release of bile and stimulates relaxation of the sphincter of Oddi allowing bile to flow into the duodenum.
Enterohepatic circulation
The enterohepatic circulation is important for the reabsorption of bile salts that are secreted into bile.
Approximately 95% of bile salts are reabsorbed in the terminal ileum and passed back to the liver via the portal system.
Disruption to the enterohepatic circulation (e.g. Crohn’s disease) can lead to the development of gallstones. The imbalance between cholesterol, bile salts and lecithin can lead gallstone formation.

